|
Separation of male and female flowers and wind-blown pollen allows open-pollinated corn plants to gain diversity of genetics. Creation of consistent characters in a hybrid, however, requires creation of pure inbreds. This process of self processing carries with it some interesting risks as well, including expression of some negative characters some of which account for the reduced size of inbreds.
Intentionally self-pollinating corn caused increasing revelation of mutations, some of which had negative effects on plant development. Corn has at least 32000 genes. As a diploid plant, having 2 copies of each chromosome, if the mutation only occurred on one DNA code in one of the pair, there would be an opportunity for the dominant form on the other member of the pair to cover up the deficiency. Self-pollination offered the chance (25%) of the new seed would be homozygous recessive and result in the expression of the mutation. What could be the effects of these mutations? Perhaps it is beneficial to our needs, like the sugary gene that gives us sweet corn, but homozygous recessives could also affect root growth, or vascular tissue size, photosynthetic rates or movement of sugars to the grain. Whereas keeping desirable ears for corn shows essentially maintained some heterosis within a variety, the open pollination nature of corn also allowed some selfing and therefore more homozygous genes for some negative characters in terms of grain production. The move to inbred development was increasing the probability of homozygous negative recessives not being covered with dominant forms of the gene within the inbred. Not only could mutations be occurring during meiosis in the selfing generation but also those accumulated within the initial breeding population, whether from an F1 cross of two inbreds or from an established variety. Inbreds inevitably will accumulate an expression of mutations that have a negative effect on grain production performance. However, with the right combination, each hybrid parent will have a dominant form of enough negative recessive forms to overcome the major disadvantages of the other parent. Experience has shown that inbreds developed from Stiff Stalk Synthetic background have a high probability of showing hybrid vigor when crossed with inbreds developed from Lancaster Sure Crop background. However, corn breeders also are aware that the combinations for inbreds from these two backgrounds that show adequate performance to be commercially competitive are not frequent. This is apparently because of the frequency of negative recessives that become homozygous during the selfing process. Given the large number of genes in corn this does seem reasonable. Although a few inbreds may combine with a few inbreds of the other heterotic group to produce close to superior commercial hybrids, and thus be considered a general combiner, ultimately there will be one specific combination giving a truly superior hybrid, at least in the environments used for that hybrid. One such hybrid 30-40 years ago was B73 X Mo17. Either of these inbreds would successfully combine with other inbreds but this specific combination was dominant in the Midwest with plant densities used during that period. However, this hybrid would also have stalk rot problems when stressed with higher plant densities or photosynthetic stress (cloudiness) more frequent in the Eastern corn belt. It is as if each combination of inbreds does not sufficiently cover all the pertinent negative genes, at least for the ultimate performance under all commercial environments but some specific crosses may be acceptable under some environments. Single cross hybrids have grain production advantages because of specific combination of the genetics of the two inbred parents plus uniform plant height and pollination timing.
Essential to reproducing identical hybrid seed corn is use of homozygous parent seed. This requires inbred seed production isolated from outside pollen, which is no easy task given that corn’s basic advantage for genetic variability is its tendency for cross fertilization. Even if the parent seed is relatively clean of contaminants, the problem continues into hybrid seed production. Traditionally, outcrosses have been often defined as plants taller than the majority of plants in a field. This has been misleading. Several years ago, we intentionally made outcrosses by pollinating a female hybrid parent with pollen from hybrid plants. That seed was planted along with the correct hybrid seed for comparison. Some of the outcrosses were taller than the correct hybrid and some were much shorter. Outcross plants varied greatly in timing of pollen and silk production as well and, significantly, were all different from each other. This is consistent with what we see in seedlings (Seedling Growouts®), each outcross plant is different from the correct hybrid and different from each other. Outcrosses that originate with pollen coming from hybrid plants are different because that pollen is the result of meiosis in the hybrid plant. Meiosis results in only one member of each of the 10 pairs of chromosomes to be represented. Minimum number of possible combinations of genetics is 2log10 or 1024 combinations of chromosomes among the pollen grain. Actually, more than that are probable because of chromosome crossovers and mutations that also occur during meiosis. Evaluation based upon taller plants is also influenced by plant height of the correct hybrid versus that of contaminating hybrid. If the correct hybrid is produced by a tall female inbred crossed with a short male inbred, it is more likely that pollen from a tall commercial hybrid will cause more ‘talls’. Even in that case, the pollen that happen to include more of the contaminating hybrid male chromosomes are likely to be smaller than the correct hybrid. Hybrid seed with less than 1% outcrosses, tall or short, have an insignificant effect on yield. Outcrosses have become more important to those with GMO interests. If the contaminating hybrid included a gene for a GMO from one of its parents, half of the resulting pollen would include that gene. If the seed production field was intended to be non-GMO, then half of the outcrosses would include that gene. If the contaminating pollen included multiple traits on separate chromosomes then the math becomes more complicated, with each of those chromosomes having a 50% chance of being included in each pollen grain. If the trait genes were closely linked on the same chromosome then they likely would be carried together but not always because some of the linkages may break. If the intended hybrid has the gene on the male parent only, then some or all the outcrosses from a non-trait hybrid would lack the gene. Selfing of the female parent in the seed production field would also lack the gene. These cases are most relevant to herbicide resistant traits and for that reason it is preferred that these genes are in the female parent genetics. Grain production involves similar dynamics except that hybrid plants tend to produce sufficient pollen to reduce the probability of contamination. It does present apprehension about strict non-gmo rules, including sampling and testing procedures. Separation of male and female flowers in corn and natural aerial distribution of pollen allowed corn to have broad genetics during the domestication of the species and adaptation of many environments. It also carries with that a small problem when we try to maintain specific trait purity such as in specific grain characteristics (ie. white or amylose corn) or GMO traits. Genetic variability in corn allows for selection of preferred genetics of human’s use. However, efficiency of production favors each plant in a field to have same maturity and other genetic characters. This is the great advantage of single cross hybrids.
Seed producers attempt to make genetically pure hybrid seed by using homozygous parents. This is not easy. Inbreeding theoretically makes gene have the identical code on the other member of the chromosome pair and thus be homozygous dominant or homozygous recessive. Small mutations can occur along the way of developing these inbreds that can interfere with the process. A larger potential source of problems can come as the inbred in increased when unintended pollen invades the seed increase field. Corn pollen viability can last only a few hours if the weather permits. Most corn pollen falls within a few feet to a few hundred feet, depending upon wind, although there is evidence of viable corn pollen found ½ mile from the source if wind and humidity is favorable. Fields for increasing parent seed inbreds are consequently isolated from other corn fields at long distances to minimize the contamination. Hybrid production using pure inbred parents also must struggle with the same problem but with a few more dynamics. Both parents are inspected visibly looking for off type plants in the male and female rows, removing those that are obvious. Most off types will show some hybrid vigor over the inbreds and thus can be easily identified and removed before pollination. Hybrid seed is produced by preventing pollen from the designated female plants by either removing the tassels or use of male sterile female inbreds. The intent of the seed producer is to have adequate pollen from the male parent to cover all exposed female silk as they emerge. The small presence of foreign pollen in the air at the same time always causes the potential for contamination, making the timing of male parent pollen production essential. Environments greatly affect the success of this endeavor. Dry field conditions tend to delay silk elongation whereas it has little effect on pollen production. This can cause most male parent pollen to be released before silks are exposed. Wet field conditions allow more elongation of silks causing the risk of silks exposed before male inbred pollen is released. Exposure of silks when little intended pollen is present increases the probability of the wrong pollen landing the on the silk and thus fertilizing the egg cell in the ovule at the end of the silk. The first viable pollen grain to arrive ‘wins’! Unintended genetics in production of hybrid seed production because of potential contamination is nearly impossible to avoid. If foreign pollen comes from a commercial hybrid field, all of the off types will not be identical. Genetics of that hybrid will include a strand of DNA from that hybrid’s male parent and a strand from the female parent. With meiosis occurring in the production of the hybrids pollen resulting in a minimum of 1024 new combinations of the 10 chromosomes in the pollen, contaminating pollen leads to many new genetics in new seed. Consequently, the outcrosses among intended hybrid seed vary greatly in appearance, some being taller than the canopy and some being shorter, along with a wide range of other morphological features. Usually contamination is associated with timing and production of intended male parent pollen thus the outcrosses often are more common within a seed size with the seed lot. First silks to emerge are from the base of the ear and thus the larger sizes. Poorly pollinated ears, perhaps from those in which silks emerged after most intended male pollen was exhausted and thus more likely to be pollinated by outside pollen results in more large, round seed. Best purity usually is in those from the middle of the ear of well pollinated ears (the medium flats). Although a few contaminants in hybrids are common and have practically no effect on hybrid yield, they gain significance in special trait hybrids. Zea mays plant structure with the strong distance between the male and female flowers and the ease of pollen movement in wind allows cross pollination of among corn plants. Easy transport of seed both of its historic movement from those initial development in Mexico as well as in recent times has allowed diversity among the 25-30000 genes in the species. This phenomenon is the major contributor to corn’s success to converting solar energy for human’s use.
Each gene of corn is a long chain of nucleic acid components in which one small substitution of that DNA can affect the performance of the resulting protein. That protein may be an important contributor to some physiological event in the plants cells. Diploid plants have a code for the same gene on the other string of DNA in that chromosome. This allows compensation for potential negative affect of any mutation in a gene on one of the pair of chromosomes. Self pollinated plants have the advantage and disadvantage of eventually getting the same mutations in both members of the chromosome pairs. This can result in selection by breeders to stabilize favorable characters but the large number of genes and probability of also getting ‘negative’ genes on both members of a chromosome pair is especially high. Inbred plants have little genetic diversity among each other. This allows for retaining genetics when in a controlled field environment like a seedstock increase isolation where all plants have identical, homozygous genes. Unfortunately, some of the affect is negative affect of some genes affecting plant size and perhaps some specific traits such as resistance to a pathogen. It is the corn breeders objective to match inbreds that will have dominant genes to reduce the negative affect when crossed with a specific inbred. After identifying the pair of inbreds that make desired hybrids with maximum expression of desired traits a commercial hybrid can be produced. Controlled pollination of one parent onto the other parent is essential for resulting seed to uniformly express the hybrid genetics. Few pollen from the female plant landing on its own ear silk, will result in seed with the female inbred and not that of the hybrid genetics. Such ‘self’ will be small, unproductive plants in the hybrid field. If female silks are pollinated by the wrong plants, such as from other hybrid fields, resulting seed will not be identical to the desired hybrid. Pollen produced by hybrid plants are not genetically identical to each other and therefore such contaminating pollen on a female plant in a hybrid seed production field results in seed with wide variation. Each outcross plant from this type of contamination will be different from the desired hybrid but also from each other, varying in plant height, ear placement and size and other noticed traits. Corn diversity benefits availability of traits and needs to be carefully controlled to maximize its advantages. Zea mays is a species that has sufficient morphological distance between the male and female flowers that most pollen does not fertilize its mother plant. This character is unique from most other crop plants in which the stamens and pistils are parts of the same flower. Although other species may have other strategies to reduce self pollination, like timing, the distance between the ear shoot and the tassel plus abundance of pollen that easily floats in air encourages pollination of other corn plants rather than self pollination.
This characteristic of corn promoted significant genetic diversity ultimately resulting in ease of adaptation to a wide range of environments as humans selected the genetics suited to their needs and environments. This is diversity to us as we selected genes for less starch in sweet corn, endosperm explosiveness in popcorn, soft starch in floury corn and hard endosperm of flint corn. Corn varieties are adapted to short seasons of temperate zones and to longer seasons near the equator. Even as pathogens evolve adaptation of some corn genetically controlled resistance, genetic variability within the species is eventually identified to produce the metabolic products needed to resist the pathogen. Open pollinated varieties maintain genetic heterozygosity as meiosis after fusion of male gamete with that of the egg cell, creates new mixes of the genes. Consequently, each seed in an open pollenated corn pollinated plant when at least one of the parents is heterozygous will have some genetic variation from adjacent seeds. Corn breeders have used this principle to get new, desired genetic characters. Genetic variability is advantageous when identifying useful characters, but not when attempting to get optimum grain production. The advantage of crossing two homozygous plants of two distinct genetic backgrounds is producing predictable and consistent hybrid plants. Controlled selfed pollination for several generation ultimately results in homozygosity, with essentially each seed having identical genetics. This selfing process also creates homozygosity of some genes that have negative affects on the plants, including characters affecting seed size and volume. Identifying homozygous inbreds that essentially overcome these negative genes of another hybrid parent is the plant breeders job for selecting potential high performing hybrids. Cross pollination characteristic of corn has allowed genetic variability and human effort to select and self pollinate to create inbreds and then crosses to make specific hybrids, allowing this species to be a major international crop. As many humans on earth face current interactions with a deadly virus, it is a reminder of significance of genetics and environmental interactions that affects our lives. Our genetic complexity affects each individual’s ability to defend against the Covid-19 virus but even that is qualified by the multiple individuals medical conditions because of past ‘environments’ and probably genetic changes in the relatively simple virus genes.
Human genetic diversity exists because of our evolution under multiple selection pressures of our past environments. All humans, despite evident, minor visible trait differences such as skin, hair and eye color, have other less evident genetic differences witnessed only by slight differences among an individual family. Corn has a genetic history somewhat similar to humans. Its ancestor was a related species and perhaps some crossing with another species but with human’s intervention it was moved to multiple environments and selection pressures allowing diversity of characters we could easily see. Yet, within each environment there was selection for other favored, less easily seen, characteristics. Just as with human the vast majority of genetics remain close to being identical to others of the species. Corn genetics that were best for desired traits in the season of an individual field in 2020 probably will not be the same as the best hybrid in another field nor even the best for the same farm in 2021. Small differences in 2021 environments will favor slightly different genetics affecting expression of traits not noted in 2020. Some traits observed in 2020 environments may not be expressed in all environments. We are dependent on maintaining genetic diversity within the corn species because of environmental variability. Humans also are dependent upon diversity not for those relatively simply inherited characters such as skin, hair and eye color but also for those less obvious genetics that our species successful adaptation to a forever changing environment. Success of our species, and of corn, is dependent upon genetic diversity. The biology of both species favors continual genetic interactions and diversity. 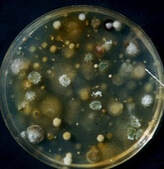 Those occupants of dying stalks, roots and ears will be there next spring as they continue to look for nutrition on organic material to digest. Living plants and animal tissue evolved with resistance to most of these organisms but we are all dependent on these organisms to release the components to be used again in the new generation.
Seeds are surrounded by many microorganisms mostly feeding on organic matter that is mostly dead. The photo, copied from our publication of Stalk Rot of Corn power point, shows the multiple colonies grown from a particle of soil in the spring of a corn field. These organisms are competing for the carbs locked up in the dead debris in the soil. Fusarium species are among these that can also penetrate some living corn roots. Fusarium is a fungus genus composed of many species. This group of fungi asexually produce spores called conidia that move in the wind. Most of these species also have a sexual stage in which the haploid hyphae fuse to form a diploid structure (ascus) in which the nuclei undergo meiosis, producing haploid ascospores, to produce more hyphae with new genetic combinations. The fact that the asexual and sexual stages are separate in time and, sometimes, location adds to the difficulty for specialists to associate the two. The ‘rules’ of naming these fungi declares that the sexual stage is primary. Consequently, on fungus commonly infecting corn seedlings, stalks and ears is known as Fusarium graminearum but also by its sexual stage name of Gibberella zeae, another common Fusarium infecting seedlings, stalks and ears is Fusarium verticillioides also known for the sexual stage name of Gibberella fujikuroi. Adding to the naming confusion, the previous name for F. verticillioides was Fusarium moniliforme. The names are confusing, but biology of these fungi is not much better. They tend to mostly be weak pathogens. They can enter weakened root and mesocotyl tissue of the germinating seed. Host resistance seems to limit the fungus ability to destroy much tissue, but it appears to simply live in the corn tissue without causing visible damage. It is not unusual to find at least a few seed germinating on a paper towel that look normal but have some hyphae of Fusarium verticillioides growth. This species frequently can be isolated from corn leaf tissue with no apparent damage. Likewise, Fusarium species are nearly always isolated from stalks of corn plants. Often, if no symptom associated with another pathogen, such as the black streaks in outer rind caused by the anthracnose fungus (Colletotrichum graminicola), we tend to call it Fusarium stalk rot. The death of the stalk mostly was a biological problem of the corn plant and fungi like Fusarium was there, among others, to assist in the digestion of dead stalk tissue. Fusarium is the name of genus of a fungus group of species that are common saprophytes and plant pathogens. Ability to kill living cells varies among Fusarium species but certainly they can feed on dead cell tissue. Among the many fungal species feeding on dead corn stalks almost always a Fusarium species will be present. If no diagnostic indicators of Gibberella, anthracnose or Diplodia are present, it becomes easy to call the disease Fusarium stalk rot.
Fusarium species that are often found in dead and live corn plant stalks initially grow between cells, producing enzymes to digest the walls of the pith cells and fibers. These walls are composed of complex cellulose compounds combined with proteins and basic carbohydrate molecules, formed by biological activity within the living cells and secreted through the cellular membranes. These compounds protect the cellular contents by structure and some active resistance products as well. Cell wall components include cellulose, arabinoxylans, hydroxycinnamates, pectins, glycoproteins and lignins. Fungi initially grow between the cells, secreting enzymes to digest the cell wall structures. Meanwhile living corn cells produce compounds such as Jasmonic acid, abscisic acid and salicylic acid to limit the growth of the Fusarium species. This battle begins with the first incursion of the fungus, perhaps with injury to the seedlings or perhaps with corn borer. Artificially infecting the stalk with any of the fungi associated with corn stalk rot shortly after pollination usually shows infection limited to area near to point of inoculation. As the tissue matures the fungi seem to more rapidly spread, digesting more cell walls. Breaking down the complex compounds allows the fungus to utilize more simple carbohydrates for their own metabolism. Digestion of the cell wall components weakens the strength of the stalk beyond the sudden decrease of strength that occurred with the plant wilted after root rot. The wilting caused the pith cells to withdraw from the rind, effectively changing the structure from a rod to a tube. This alone is believed to reduce strength by one-third. Corn hybrids that tend to construct more and stronger cells in the rind can have less lodging but the physical change from a rod to tube is difficult to match by cell wall strength alone. Maintaining sturdy stalks until harvest is a balance among producing sufficient carbohydrates for filling all kernels, avoiding wilt by delaying death of root cells and maintaining stalk cells alive to fend off potential pathogens such as Fusarium Less common in the USA corn belt but occasionally regionally destructive is Head Smut. It is not easily distinguished from common smut, but it has distinctive characters and resistance is more common. This smut-causing fungus is Sphacelotheca reiliana. This smut shows mostly in the complete destruction of the ear and the tassel of corn. This species also attacks sorghum, Johnson grass and Teosinte. The diploid teliospores of this fungus, shed from the smutted ears and tassel remain dormant in the soils for many years but when activated by the root exudates from susceptible corn seedling, the spore undergoes meiosis, producing short hyphae and then haploid basidiospores. These spores germinate to form hyphae that fuse with compatible haploid hyphae, resulting the cells with two haploid nuclei. From this point the fungus infects root cells, apparently growing between the cell wall and the cell membrane of the affected cell.
Absorbing the nutrients from the infected, live cell, the fungus continues growing cell to cell towards the meristems of the seedling. From there it continues to advance without killing corn cells as they undergo cell division and eventually producing the ear and tassel. Disease symptoms from the infection are absent as the fungus absorbs nutrients but does not kill tissue nor penetrate the cell membranes. After the host plant’s meristematic cells at the upper tip of the stalk and ear change to producing pollen mother cells and ovules, the fungal cells also change. The two compatible nuclei in each fungal cell fuse to form diploid single cells. These fungal diploid cells fill and destroy the host cells in the tassel and ears before pollination. The individual, diploid fungal cells develop thick walls resulting in the first sign of the disease as a loose powder forms instead of ears on the corn plants. These teliospores are then spread by the wind, the thick wall giving them survival until another host plant is found for repetition of the cycle. Fortunately, high levels of resistance to head smut is more common than susceptibility. Some of the resistance is thought to be from differences in the root exudates in the initial induction of teliospore germination. Some comes from the successful recognition of the initial invasion, triggering the response by the host of producing enzymes to keep the pathogen from reaching the meristems. Perhaps it involves the speed of the plants cell divisions, escaping the fungus while infected cells produce anti-fungal chemistry. Regardless of the method, most corn hybrids are reasonably resistant. The problem historically has been with popular, susceptible hybrids being planted for consecutive years, allowing an increasing concentration of teliospores, until the right environment such as warm, light soils at time of corn seed germination results in noticeable high infection rate. Switching to more resistant hybrids in future seasons reduces the occurrence of head smut. Biology of corn includes the biology of pathogens. |
About Corn JournalThe purpose of this blog is to share perspectives of the biology of corn, its seed and diseases in a mix of technical and not so technical terms with all who are interested in this major crop. With more technical references to any of the topics easily available on the web with a search of key words, the blog will rarely cite references but will attempt to be accurate. Comments are welcome but will be screened before publishing. Comments and questions directed to the author by emails are encouraged.
Archives
December 2021
Categories
|
 RSS Feed
RSS Feed