|
Humans and plants have similarities in fighting pathogens. Structural characters such as skin on animals and tight epidermal cells on plant leaves prevent invasion by most micro-organisms capable of destroying internal tissues rich in nutrition for them. But evolution favors some organisms able to avoid this outer defense system, requiring the host to detect their presence and turn on resistance systems.
Corn leaf epidermal cells are tightly connected except for the stomata. The vast majority of microbes surrounding corn plants cannot penetrate the plants. The few that do have the capability, perhaps by enzymatically drilling through the epidermal cells to enter the leaf tissue, set off the alarm. In some cases, the plant’s first response is increasing the production of salicylic acid in the area of the invasion. This turns on the genes for production of the protein, often an enzyme, with the capacity of stopping the pathogen from spreading. This final product may be effective against several potential pathogens or specific to one species. There are genetics, of course, behind the chemical responses to attacks by pathogens. The genetics must relate to detection of an attack, perhaps detection of the specific pathogen, production of a general or specific anti-pathogen material and speed of the response. Although all resistance to corn diseases involves multiple genes affecting the biology for the processes mentioned above, in many diseases there is a single corn gene that has a drastic, critical affect on the pathogen. This is called qualitative resistance. In the case of rust diseases, the pathogen is killed almost as quickly as it invades a cell. With Exserohilum turcicum the plant with an Ht gene stops the fungus as it enters the vascular system, inhibiting the fungus from producing usual northern leaf blight lesions and spores to further spread the disease. Qualitative resistance is convenient for the corn breeder to select during the breeding process but, unfortunately, nearly always the population of the pathogen includes individuals with a single gene that produces a product to overcome the resistance product. Eventually, natural selection results in increasing presence of those pathogens with this gene and the the corn single gene is less useful. The more stable resistance involves strengthening the detection, and the speed and concentration of the anti-pathogen materials. This is called quantitative resistance. It is controlled by several genes and therefore more difficult to identify by breeders. It is usually reflected by fewer and smaller lesions but assurance of exposure to the pathogen comparable to potential for the commercial life of the hybrid is not simple. Most diseases are heavily influenced by environments which vary by location and season. Most susceptible genetics are eliminated in breeding nurseries, but we do get surprised occasionally either with increased intensity of a disease or new pathogen changes. Humans are witnessing the affects of genetics of a virus affecting human behavior everywhere. Dynamics of the COVID-19 and people interactions will have some permanent affects, some of which are not yet clear. Biology, affected by genetics, in plants and their pathogens have similar long-term interactions. We witnessed this in the outbreak of the Southern Corn Leaf Blight in 1969 and 1970. That phenomenon was blogged in Corn Journal issue 129/2019.
Race t of Helminthosporium maydis(Bipolaris maydis) (Cochliobolus heterostrophus) spread across most corn growing areas in USA and elsewhere in 1970. The traditional version, race 0, of this pathogen was common in the Southeastern USA where temperatures and humidity favored the biology of the fungus. A related fungus Helminthosporium carbonum (Bipolaris zeicola) (Cochliobolus carbonum) was a common pathogen of corn but tended to be more frequent in the northern part of the US corn belt. The summer of 1970 not only featured epidemics of race T of H. maydis but spread of this pathogen to much of northern corn belt. This allowed the co-mingling of the two species. The two species were distinguished by microscopic examination of their conidia, the asexually produced spores associated with spread of these fungi. H. maydis spores were consistently curved and appeared to be gray when viewed with a light microscope. H. carbonum conidia were darker in pigment and mostly straight. Both species had shown to have similar sexual reproduction structures and to have distinct sexual mating types. Seed companies, including the one that I had just joined, were checking their inbreds and hybrids in the summer of 1972 to make sure there was no remnant susceptibility left among their materials. I was surprised to find a wide range of shapes and sizes of lesions naturally occurring among materials that looked like southern corn leaf blight in our central Illinois nursery. Examining the spores under microscope showed a range of spore shapes intermediate to H. maydis and H. carbonum. Other pathologists found the same thing. It had been shown previously that these two species could cross in lab experiments and now it appeared that the wide-spread distribution of H. maydis into regions where H. carbonum was common allowed multiple opportunities for sexual crosses between the two. This apparently accounted for the range of conidia morphology seen in the summer of 1972. The resulting population of these multiple crosses further sorted in virulence on corn. H. carbonum already had been found, with one race (race 1) to produce a toxin affecting a few inbreds homozygous recessive to the toxin. Another group of this pathogen appeared to mildly pathogenic on corn leaves was defined as Race 2. After 1972, some inbreds and hybrids were found to be susceptible to Race 3, resulting in distinctive long, narrow lesions. In 1980, inbreds with B73 backgrounds, commonly used as female parents in seed production fields, were infected with a distinct race 4 of H. carbonum. Apparently, the crosses of the two related fungal species resulted in new genetic combinations. This event occurring 50 years ago not only changed corn breeders wariness of corn’s disease vulnerability but also all involved with corn to be constantly observing for subtle changes in pathogen-host problems. Biology of one affects biology of others. Corn nucleus includes 30000-40000 genes in its chromosomes. These 10 chromosomes are sorted and combined with the chromosomes of the other parent during pollination, both parents contributing to the hybrid. Cell organelles, such as plastids and mitochondria, have up to 100 genes in each small structure but these are not contributed by both parents during pollination. All organelle DNA is contributed by the egg cell.
Small mutations in the in mitochondria of the female parent can result in drastic affects on the plant. Such mutations resulted in cytoplasmic male sterility in CMS corn because in reduced the energy within pollen producing cells to make fertile pollen. This happens in T, C and S male sterility. Mitochondria function however is not completely dependent upon its own DNA, however, as nuclear genes in the cell also affect mitochondria function. Fertile pollen can be restored by cell nuclear genes such as the Rf1 and Rf2 that make T cytoplasm corn fertile. Similar fertility restoration for C and S cytoplasm sterility and be restored with presence of specific genes in the nuclei of the cells. Plastids in cells function locations of starch storage and, in leaves, as the site of photosynthesis. These chloroplasts also have their own DNA with about 104 genes. Corn is among the few C4 plants the process photosynthesis in two distinct types of chloroplasts. Those in the main leaf cells do the initial process but final steps are carried out in chloroplasts in the bundle cells surrounding the cells. This is reviewed in Corn Journal blog of 7/14/2020 and 7/1/2020. Corn hybrid genetics are mostly affected by the combination of specific nuclear genetic combinations of the two parent inbreds. Because only the female parent contributes the genetics of mitochondria and plastids choosing which parent is to be the female can have a drastic affect on some plant performance factors. This can be dramatic with some parents having reduced germinations. It is complicated and wonderful! Corn like other plants and animals have some DNA in cell organelles outside of the chromosomes in the cell nucleus. Mitochondria are the site in the cell where carbohydrates are transformed into useable chemical form (ATP) for most energy use in the cell. It is an energy bound organelle in which these reactions occur. This Corn Journal blog in 2017 describes a dramatic interaction of corn mitochondria mutation and a devastating pathogen.
All living cells of plants and animals have mitochondria, organelles that convert carbohydrates into the useful form of energy that drives synthesis of metabolites in cells. Mitochondria are believed to be descendants of bacteria that became symbiotic with cells in the early evolution of most living forms. They retained their own DNA, are transferred to the next generation only in eggs cell and not sperm. They replicate within cells, but the host cells have some control on the rate of replication. Energy conversion in mitochondria occurs on their folded membranes in a series of chemical reactions. Regions of the plant undergoing rapid cell duplication have more mitochondria. This includes the tassel cells of a corn plant. The pollen mother cells in that region undergo meiosis and duplication, driven partly by the energy conversion by concentration of mitochondria in those mother cells. A small defect in mitochondrial DNA of an inbred caused a defective membrane product in those mitochondria resulting in incomplete development of pollen. This was found in a corn breeding program in Texas. As the inheritance of this condition was known to be only transmitted independent of nuclear DNA, it was called Texas male sterile cytoplasm. It became a useful tool to corn hybrid seed production because it was easily transferred in breeding programs to the female parent of a hybrid, and thus avoiding manual removal of tassels in seed production fields. Use of T male-sterile cytoplasm became common in the worldwide corn in the 1960’s. It was noted in the Philippines in 1961, that a fungal pathogen, then known as Helminthosporium maydis, was especially aggressive on several hybrids with T cytoplasm. Despite a few scattered reports elsewhere it was not until 1969 that the connection between increased occurrence of this disease and T cytoplasm became alarming. Majority of seed produced for 1970 corn season had T cytoplasm, the main exceptions being new hybrids in which the conversion to sterility of the female parents was incomplete. Although the pathogen was normally found in the southern half of the corn belt, and adequately controlled by products of nuclear DNA genes, this disease was found highly destructive in northern corn belt areas as well. A race of the fungus (now named Bipolaris maydis and by its sexual stage Cochliobolus heterostrophus) called race T, produces a toxin that causes death to cells with mitochondria having the DNA with the defect associated with T male sterility. All cells of the corn plant with these defective mitochondria were vulnerable to the fungus. This included the cells in developing seed resulting in diseased stored grain as well as overwintering leaves and stalks. Normal resistance mechanisms to the pathogen were ineffective because the toxin destroyed these defective mitochondria. As the relationship with T cytoplasm was realized, seed companies worked to change, and within a few years, the disease subsided back to its normal distribution. It was a new learning experience of interaction of corn and pathogen biology. Fungi, bacteria and viruses have constant genetic mutations and evolution favor those that adapt to potential food sources. This pathogen diversity must be matched with diversity in corn to avoid crop losses. The list of new, unexpected occurrences in USA and internationally seem to occur every few years. Probably we should not be surprised. As we attempt to improve the hybrids for yield, standability, performance under changing environments we can also inadvertently and unknowingly include genes for susceptibility. Especially vulnerable are genes that allow the recognition of the invader and therefore quick defense response. Furthermore, a corn hybrid that may work well, and not express susceptibility to local pathogens, but when moved to another environment, a pathogen reacts differently.
Microbes also have genes that vary from mutations and sexual recombination. Rapid production of huge numbers of spores, ability to infect multiple hosts often near corn field, and broad, widespread populations of these pathogens is their strength. New variants of the pathogen, adapted to at least a few current hybrids with higher level of pathogenicity, must initially show only in isolated spots in corn fields and easily overlooked. History nearly all ‘new’ races in the USA were not noted until they were widespread. My guess is the genetic variant allowing for the new race (pathotype) was infrequent but present in the pathogen population for some time but was not recognized until damage was common. The more recent occurrence of the bacterial streak of corn, caused by Xanthomonas varicola pv. vasculorum, previously known only in South Africa but within only a few years it was been identified in several US states and Argentina. Was it here for a long time, or distributed by seed or even grain debris? Perhaps it has been a long-time pathogen of other grasses. Or perhaps is a mutant of a related bacterium and we inadvertently selected for susceptibility in corn. It is doubtful that the battle between new pathogen variants and corn will end. Our best protection must come from careful observations in corn fields and submitting suspicious samples to appropriate specialists for identification. Corn genetic diversity has always allowed selection of resistance, but it does take a few years to implement the hybrid seed production before serious damage to the crop. The fun never stops! Constant interactions with the corn plant biology, affected by its genetics, and pathogen biology, affected by its genetics, requires us to carefully observe potential changes. Each participant in these interactions has potential for reducing our final product from the corn crop.
Bacterial pathogenesis of corn differs from fungal pathogenesis mostly because of biological differences in the two types of organisms. Bacteria are single-celled organisms with a single chromosome without a nucleus in its cytoplasm. Cell division in bacteria results in a new, distinct individual cell, whereas fungal cells divide to form a filament, frequently maintaining cell to cell communication and specialization. Bacteria cell single chromosome simply replicates, then separates as a cell wall divides the cell into two halves, each with its own single chromosome. Fungal chromosomes are enclosed in a nuclear membrane, divide by mitosis during cell division but also can undergo meiosis forming single strands of the chromosome and thus exist as haploids until the nuclei fuse to form diploids. These structural and functional differences between bacteria and fungi results in different ‘strategies’ concerning adaptations of these organisms to changing environments. Mutations occur in both, but those in the single bacterium chromosome immediately results in a changed new individual bacterium. Mutation in fungal DNA may not take effect until recombination to form the homozygous recessive. Bacterial biology allowing rapid reproduction of the new forms allows adjustment to new hosts. Bacteria are ubiquitous, distributed by wind, water, insects and animals. Rapid cell reproduction allows quick spread when in a nutritional environment. Despite their widespread presence, most species cannot infect living corn leaves. Leaves have evolved epidermal cells tightly held together and with a waxy covering, repelling water and prohibiting bacterial infection. Pathogenic fungi establish a multicellular mat (appresorium) from which it produces a penetration peg that enzymatically drills through the wax and epidermal walls to enter the leaf. Bacteria cells can enter through open stomata-some even have flagella to help swim to the stomata. Plant resistance systems can produce antibacterial fumigants to inhibit the bacteria from replicating and there is some evidence stomata close when faced with bacteria invasion. Small size and few visible structural features has made it difficult to identify a bacterium causing a plant disease. Although some disease symptoms do become associated with specific species, confirmation usually requires culturing and chemical tests. It often takes a simple mutation in a potential pathogen, or in the corn host, for a relatively minor disease to become major. Good for everyone in corn production to be aware as the crop grows. Virus genetics are probably the simplest of pathogens, some not even bothering with DNA but using mRNA to directly cause the host cell to provide the more complex process of DNA code as well as the ribosomes to produce essential proteins for the pathogen. Fungal pathogens of corn have more complex genetic systems than viruses and corn.
Corn reproduction is relatively easy to understand when compared to most corn pathogens. Most corn cells have a single nucleus with 10 pairs of chromosomes with only the exceptions of the triploid cells of the endosperm and the haploid cells in the pollen and egg cells. The hyphae of most corn leaf pathogens are filaments composed of multiple cells separated by cell walls called septa that have pores allowing exchange of cytoplasm and even nuclei. The nuclei are composed of a single set of chromosomes. Furthermore, a single cell of the fungus causing northern leaf blight may have up to 30 nuclei, all haploid. Genetics of diploid cells are affected by both sets of the pair, generally with the dominant form of a gene on one set of the chromosomes resulting in the amino acid components of the protein and therefor the trait expressed. Recessive genes get expressed only when both sets have the same recessive i.e., homozygous recessive for that gene. Mutations resulting in new recessive genes are often ineffective because of the dominant form in the other member of the paired chromosome. In a haploid nucleus, there is only one form of a gene, allowing a recessive gene to be fully expressed. Resistance, especially those with single gene inheritance, in corn to a pathogen such as Exserohilum turcicum, cause of northern leaf blight, is initiated when the host cells recognize the presence a pathogens product (effector). A mutation in the pathogen’s gene for this effector, negates the plants ability to recognize the pathogen and therefore this single gene for resistance is not initiated. This is the weakness of the single gene system for resistance to any pathogen. Fungal pathogens have the statistical advantage of producing new races because of the haploid nuclei in most hyphae. For example, if a homozygous recessive form is crossed with a homozygous dominant, the first generation of a diploid organism would only produce the dominant gene product. In the haploid hyphae, half of the resulting cells would produce the recessively inherited product. This explains why some pathogens can successfully overcome the single gene type of resistance. Being able to asexually reproduce with thousands of spores with haploid nuclei from the successful pathogen allows quick spread of a new genetic type. Plant and animal genetics involve a complex involvement of DNA (deoxyribonucleic acid) with long chains of long chains of nucleic acids arranged in sets of 4. These chains are arranged as chromosomes and enclosed in a membrane. The DNA also codes for a related string of nucleic acids called RNA (ribonucleic acid). When a segment of the DNA is signaled to be ‘read’, that string of nucleic acids is transferred to messenger RNA. This mRNA moves through the nuclear membrane to a ribosome in the cell. Each set of 4 nucleic acids link specific amino acids to produce a specific protein. Thus, the DNA-RNA codes result in specific proteins produced in animal and plant cells. Many of these proteins function as enzymes for basic metabolism, ultimately resulting in structure and functions of plants and animals.
Viruses have simplified the process. Some have DNA and some only have RNA. They use the hosts cell’s ribosome for the protein production needed for the virus membrane. Current human epidemic caused by a coronavirus has its genetic code in mRNA. This sufficient to cause the host cell ribosomes to produce the few proteins needed for replication and further infection by the virus. One of those protein forms the ‘spike’ allowing the virus to penetrate the cell membrane. Viruses, such as this one, have self replicating RNA, simplifying the duplication process. Viruses causing corn diseases have either RNA or DNA codes for duplication within host cells. Most appear to be RNA only genetic codes but a few, such as Maize Streak virus, has its genetic code in the form of DNA. Most corn virus require a vector to penetrate the plant cell walls, but after entering, they use the host cell’s cytoplasm to replicate. Host resistance depends upon successfully producing ‘antibodies’ to inhibit this replication process. Let’s hope that the recently revealed vaccines for current human coronavirus that apparently involve a segment of the mRNA will successfully halt the ability of the virus to replicate. tEach seed to be planted next year will have a full set of the hybrid genes located in each cell of the embryo. Natural drying of seed in the field allows a temporary delay in cellular activity in corn seed about 40 days after pollination. Respiration and other metabolism in the embryo cells slow, preventing germination. Preservation of seed viability for the next season requires continuation of the seed drying to 14-7% to prevent faster metabolism. Corn seed producers control this drying process, carefully avoiding higher temperatures that could destroy the cellular physiology but yet quickly dry the harvested seed. This is mostly done by using fans to remove the released moisture from environment surrounding the seed.
Drying speed is significant because moisture levels between 32% and 15% allow some degradation of cellular membranes but without the adequate replacement that occurs in actively growing plants. This ageing process shortens the eventual viability of the seed. Multiple studies have shown that seed respiration rates are affected by storage conditions and that these eventually affect germination percentage and seedling growth rate. Genetics also enter into these factors. Pericarp wall density affects the ease of moisture movement into the embryo. Some corn genotypes are more vulnerable to uptake of atmospheric moisture than others. Pericarp tissue is part of the female parent of the corn kernel and therefore vulnerability to seed germination deterioration due to moisture during storage is often associated with the hybrid female parent. Two inbreds may combine to give identical hybrid parent characteristics, but one of the inbreds may be superior for germination preservation when used as the female parent. Successful preservation of high germination rates in the spring are dependent on multiple factors including stresses during seed development, drying conditions and genetics of parent seed. As the Corn harvest season draws towards a close, once again a corn grower witnesses the variability within and among individual fields. Differences in soil types, crop history and perhaps even rainfall distribution affected corn hybrids’ grain yield and stalk performance. Genetic diversity among corn hybrids provide opportunity to adjust with choices among next year’s hybrids although weather for the next season is not easily predicted.
Zea mays features tremendous genetic diversity due to its cross-pollination biology and humans involvement of exposing it to multiple environments throughout its relatively short history. The large number of genes on only 10 chromosomes allows fairly rapid expression of mutations. Hybrids with deeper roots allow water absorption in sandy soils while those with more branching of roots near the surface are better adapted to soils with lots of organic matter from the previous year crop. If next year’s weather tends to be dry, especially during flowering time, the deeper root hybrid may be favored. If it is wet during that critical time period, the shallow root type may do better in grain yield and stalk quality. Mid season wind pressure may favor the hybrid with roots with more branching. Previous crop environments may influence need for resistance to some leaf diseases. Corn breeders attempt to select away from extreme susceptibility to pathogens such as Exserohilum (Helminthosporium) turcicum, cause of northern leaf blight, and Cercospora zeae-maydis, cause of gray leafspot, but genetic variability among pathogens and weather patterns influence the ultimate threat to the crop. Genetic variability among corn hybrids and within potential pathogens will always present changes in the affect of corn diseases on performance. We are dependent upon researcher in both genetics of both host and pathogen to monitor these possibilities. We are also dependent upon growers to access all agronomy knowledge, make the most economic and technical decisions in choosing hybrids for each field for next season-and then roll the dice! Seed producers attempt to make genetically pure hybrid seed by using homozygous parents. This is not easy. Inbreeding theoretically makes genes with the identical code on the other member of the chromosome pair and thus be homozygous dominant or homozygous recessive. Small mutations can occur along the way of developing these inbreds that can interfere with the process. A larger potential source of problems can come as the inbred in increased when unintended pollen invades the seed increase field.
Corn pollen viability can last only a few hours if the weather permits. Most corn pollen falls within a few feet to a few hundred feet, depending upon wind, although there is evidence of viable corn pollen found ½ mile from the source if wind and humidity is favorable. Fields for increasing parent seed inbreds are consequently isolated from other corn fields at long distances to minimize the contamination. Hybrid production using pure inbred parents also must struggle with the same problem but with a few more dynamics. Both parents are inspected visibly looking for off type plants in the male and female rows, removing those that are obvious. Most off types will show some hybrid vigor over the inbreds and thus can be easily identified and removed before pollination. Hybrid seed is produced by preventing pollen from the designated female plants by either removing the tassels or use of male sterile female inbreds. The intent of the seed producer is to have adequate pollen from the male parent to cover all exposed female silk as they emerge. The small presence of foreign pollen in the air at the same time always causes the potential for contamination, making the timing of male parent pollen production essential. Environments greatly affect the success of this endeavor. Dry field conditions tend to delay silk elongation whereas it has little effect on pollen production. This can cause most male parent pollen to be released before silks are exposed. Wet field conditions allow more elongation of silks causing the risk of silks exposed before male inbred pollen is released. Exposure of silks when little intended pollen is present increases the probability of the wrong pollen landing the on the silk and thus fertilizing the egg cell in the ovule at the end of the silk. The first viable pollen grain to arrive ‘wins’! Unintended genetics in production of hybrid seed production because of potential contamination is nearly impossible to avoid. If foreign pollen comes from a commercial hybrid field, all of the off types will not be identical. Genetics of that hybrid will include a strand of DNA from that hybrid’s male parent and a strand from the female parent. With meiosis occurring in the production of the hybrids pollen resulting in a minimum of 1024 new combinations of the 10 chromosomes in the pollen, contaminating pollen leads to many new genetics in new seed. Consequently, the outcrosses among intended hybrid seed vary greatly in appearance, some being taller than the canopy and some being shorter, along with a wide range of other morphological features. Usually contamination is associated with timing and production of intended male parent pollen thus the outcrosses often are more common within a seed size with the seed lot. First silks to emerge are from the base of the ear and thus the larger sizes. Poorly pollinated ears, perhaps from those in which silks emerged after most intended male pollen was exhausted and thus more likely to be pollinated by outside pollen results in more large, round seed. Best purity usually is in those from the middle of the ear of well pollinated ears (the medium flats). Although a few contaminants in hybrids are common and have practically no effect on hybrid yield, they gain significance in special trait hybrids. It does take special effort to produce pure hybrid seed. Observations of corn hybrids and inbreds at flowering stage and later allows distinction of genetic differences, especially if several varieties are planted in unform conditions. Usually plant height differences are clear. Close observation of ear characters also allows distinguishing between varieties especially when grown near each other with same micro-environment. Closer observation of the plants for other morphological differences such as tassel and leaf shape and size also can allow distinguishing genetic differences, although can these can be vulnerable to confusion with the differences caused by some virus infections and poor seedling emergence.
Inbreds and hybrids differ from each other in many other genes than only those observed in mature plants. Close observation of plants at all stages show other leaf character differences that becomes obvious when plants are in identical environments. Multiple leaf character differences become obvious when observed by experienced researchers. Pure single cross hybrid seed planted in controlled soil and temperature environment will be identical to each other for many leaf characters. Modified single cross hybrids, such as when the female parent is a cross of two related inbreds, will be reflected in seedlings with slight morphological differences. Outcross plants caused by pollen blown into the hybrid seed production field will show multiple character differences among the seedlings. A seed mix accident in which two hybrids in the same sample will show two distinct seedling plant types. This is also true where one of the inbred parents was mixed in the seed production field or if contaminated with the male parent from an adjacent field. Accident selfing problems in the seed field results in some plants not only with distinct seedling morphology, and usually smaller than the hybrid seedlings. These ‘selfs’ are also morphologically identical to each other. If compared to a sample of the female parent, these plants can be confirmed as ‘selfs’. Professional Seed Research, Inc. has used this information and experience to evaluate purity of hybrid and inbred seed for seed companies for more than 30 years (Seedling Growout® test). 400 seed are planted adjacent to each other in uniform environment with natural light. Experienced researchers carefully observe each plant for morphological differences to identify those that are unlike the others. If all the offtypes differ from each other, it is assumed that they are from contamination from outside hybrid fields. If identical to each other then probably the source is a seed mix or possibly contamination from an adjacent seed field. If the plants are smaller for most leaf characters yet distinct from most plants and identical to each other, they are assumed to be selfs. This method of evaluation of purity of seed samples allows for results within 2 weeks of planting, and larger samples (400 seed) than most other grow out methods. It has been used with temperate and tropical hybrids. Corn’s diverse genetics are expressed at all stages of the plant’s development. Separation of male and female flowers and wind-blown pollen allows open-pollinated corn plants to gain diversity of genetics. Creation of consistent characters in a hybrid, however, requires creation of pure inbreds. This process of self processing carries with it some interesting risks as well, including expression of some negative characters some of which account for the reduced size of inbreds.
Intentionally self-pollinating corn caused increasing revelation of mutations, some of which had negative effects on plant development. Corn has at least 32000 genes. As a diploid plant, having 2 copies of each chromosome, if the mutation only occurred on one DNA code in one of the pair, there would be an opportunity for the dominant form on the other member of the pair to cover up the deficiency. Self-pollination offered the chance (25%) of the new seed would be homozygous recessive and result in the expression of the mutation. What could be the effects of these mutations? Perhaps it is beneficial to our needs, like the sugary gene that gives us sweet corn, but homozygous recessives could also affect root growth, or vascular tissue size, photosynthetic rates or movement of sugars to the grain. Whereas keeping desirable ears for corn shows essentially maintained some heterosis within a variety, the open pollination nature of corn also allowed some selfing and therefore more homozygous genes for some negative characters in terms of grain production. The move to inbred development was increasing the probability of homozygous negative recessives not being covered with dominant forms of the gene within the inbred. Not only could mutations be occurring during meiosis in the selfing generation but also those accumulated within the initial breeding population, whether from an F1 cross of two inbreds or from an established variety. Inbreds inevitably will accumulate an expression of mutations that have a negative effect on grain production performance. However, with the right combination, each hybrid parent will have a dominant form of enough negative recessive forms to overcome the major disadvantages of the other parent. Experience has shown that inbreds developed from Stiff Stalk Synthetic background have a high probability of showing hybrid vigor when crossed with inbreds developed from Lancaster Sure Crop background. However, corn breeders also are aware that the combinations for inbreds from these two backgrounds that show adequate performance to be commercially competitive are not frequent. This is apparently because of the frequency of negative recessives that become homozygous during the selfing process. Given the large number of genes in corn this does seem reasonable. Although a few inbreds may combine with a few inbreds of the other heterotic group to produce close to superior commercial hybrids, and thus be considered a general combiner, ultimately there will be one specific combination giving a truly superior hybrid, at least in the environments used for that hybrid. One such hybrid 30-40 years ago was B73 X Mo17. Either of these inbreds would successfully combine with other inbreds but this specific combination was dominant in the Midwest with plant densities used during that period. However, this hybrid would also have stalk rot problems when stressed with higher plant densities or photosynthetic stress (cloudiness) more frequent in the Eastern corn belt. It is as if each combination of inbreds does not sufficiently cover all the pertinent negative genes, at least for the ultimate performance under all commercial environments but some specific crosses may be acceptable under some environments. Single cross hybrids have grain production advantages because of specific combination of the genetics of the two inbred parents plus uniform plant height and pollination timing.
Essential to reproducing identical hybrid seed corn is use of homozygous parent seed. This requires inbred seed production isolated from outside pollen, which is no easy task given that corn’s basic advantage for genetic variability is its tendency for cross fertilization. Even if the parent seed is relatively clean of contaminants, the problem continues into hybrid seed production. Traditionally, outcrosses have been often defined as plants taller than the majority of plants in a field. This has been misleading. Several years ago, we intentionally made outcrosses by pollinating a female hybrid parent with pollen from hybrid plants. That seed was planted along with the correct hybrid seed for comparison. Some of the outcrosses were taller than the correct hybrid and some were much shorter. Outcross plants varied greatly in timing of pollen and silk production as well and, significantly, were all different from each other. This is consistent with what we see in seedlings (Seedling Growouts®), each outcross plant is different from the correct hybrid and different from each other. Outcrosses that originate with pollen coming from hybrid plants are different because that pollen is the result of meiosis in the hybrid plant. Meiosis results in only one member of each of the 10 pairs of chromosomes to be represented. Minimum number of possible combinations of genetics is 2log10 or 1024 combinations of chromosomes among the pollen grain. Actually, more than that are probable because of chromosome crossovers and mutations that also occur during meiosis. Evaluation based upon taller plants is also influenced by plant height of the correct hybrid versus that of contaminating hybrid. If the correct hybrid is produced by a tall female inbred crossed with a short male inbred, it is more likely that pollen from a tall commercial hybrid will cause more ‘talls’. Even in that case, the pollen that happen to include more of the contaminating hybrid male chromosomes are likely to be smaller than the correct hybrid. Hybrid seed with less than 1% outcrosses, tall or short, have an insignificant effect on yield. Outcrosses have become more important to those with GMO interests. If the contaminating hybrid included a gene for a GMO from one of its parents, half of the resulting pollen would include that gene. If the seed production field was intended to be non-GMO, then half of the outcrosses would include that gene. If the contaminating pollen included multiple traits on separate chromosomes then the math becomes more complicated, with each of those chromosomes having a 50% chance of being included in each pollen grain. If the trait genes were closely linked on the same chromosome then they likely would be carried together but not always because some of the linkages may break. If the intended hybrid has the gene on the male parent only, then some or all the outcrosses from a non-trait hybrid would lack the gene. Selfing of the female parent in the seed production field would also lack the gene. These cases are most relevant to herbicide resistant traits and for that reason it is preferred that these genes are in the female parent genetics. Grain production involves similar dynamics except that hybrid plants tend to produce sufficient pollen to reduce the probability of contamination. It does present apprehension about strict non-gmo rules, including sampling and testing procedures. Separation of male and female flowers in corn and natural aerial distribution of pollen allowed corn to have broad genetics during the domestication of the species and adaptation of many environments. It also carries with that a small problem when we try to maintain specific trait purity such as in specific grain characteristics (ie. white or amylose corn) or GMO traits. Genetic variability in corn allows for selection of preferred genetics of human’s use. However, efficiency of production favors each plant in a field to have same maturity and other genetic characters. This is the great advantage of single cross hybrids.
Seed producers attempt to make genetically pure hybrid seed by using homozygous parents. This is not easy. Inbreeding theoretically makes gene have the identical code on the other member of the chromosome pair and thus be homozygous dominant or homozygous recessive. Small mutations can occur along the way of developing these inbreds that can interfere with the process. A larger potential source of problems can come as the inbred in increased when unintended pollen invades the seed increase field. Corn pollen viability can last only a few hours if the weather permits. Most corn pollen falls within a few feet to a few hundred feet, depending upon wind, although there is evidence of viable corn pollen found ½ mile from the source if wind and humidity is favorable. Fields for increasing parent seed inbreds are consequently isolated from other corn fields at long distances to minimize the contamination. Hybrid production using pure inbred parents also must struggle with the same problem but with a few more dynamics. Both parents are inspected visibly looking for off type plants in the male and female rows, removing those that are obvious. Most off types will show some hybrid vigor over the inbreds and thus can be easily identified and removed before pollination. Hybrid seed is produced by preventing pollen from the designated female plants by either removing the tassels or use of male sterile female inbreds. The intent of the seed producer is to have adequate pollen from the male parent to cover all exposed female silk as they emerge. The small presence of foreign pollen in the air at the same time always causes the potential for contamination, making the timing of male parent pollen production essential. Environments greatly affect the success of this endeavor. Dry field conditions tend to delay silk elongation whereas it has little effect on pollen production. This can cause most male parent pollen to be released before silks are exposed. Wet field conditions allow more elongation of silks causing the risk of silks exposed before male inbred pollen is released. Exposure of silks when little intended pollen is present increases the probability of the wrong pollen landing the on the silk and thus fertilizing the egg cell in the ovule at the end of the silk. The first viable pollen grain to arrive ‘wins’! Unintended genetics in production of hybrid seed production because of potential contamination is nearly impossible to avoid. If foreign pollen comes from a commercial hybrid field, all of the off types will not be identical. Genetics of that hybrid will include a strand of DNA from that hybrid’s male parent and a strand from the female parent. With meiosis occurring in the production of the hybrids pollen resulting in a minimum of 1024 new combinations of the 10 chromosomes in the pollen, contaminating pollen leads to many new genetics in new seed. Consequently, the outcrosses among intended hybrid seed vary greatly in appearance, some being taller than the canopy and some being shorter, along with a wide range of other morphological features. Usually contamination is associated with timing and production of intended male parent pollen thus the outcrosses often are more common within a seed size with the seed lot. First silks to emerge are from the base of the ear and thus the larger sizes. Poorly pollinated ears, perhaps from those in which silks emerged after most intended male pollen was exhausted and thus more likely to be pollinated by outside pollen results in more large, round seed. Best purity usually is in those from the middle of the ear of well pollinated ears (the medium flats). Although a few contaminants in hybrids are common and have practically no effect on hybrid yield, they gain significance in special trait hybrids. Zea mays plant structure with the strong distance between the male and female flowers and the ease of pollen movement in wind allows cross pollination of among corn plants. Easy transport of seed both of its historic movement from those initial development in Mexico as well as in recent times has allowed diversity among the 25-30000 genes in the species. This phenomenon is the major contributor to corn’s success to converting solar energy for human’s use.
Each gene of corn is a long chain of nucleic acid components in which one small substitution of that DNA can affect the performance of the resulting protein. That protein may be an important contributor to some physiological event in the plants cells. Diploid plants have a code for the same gene on the other string of DNA in that chromosome. This allows compensation for potential negative affect of any mutation in a gene on one of the pair of chromosomes. Self pollinated plants have the advantage and disadvantage of eventually getting the same mutations in both members of the chromosome pairs. This can result in selection by breeders to stabilize favorable characters but the large number of genes and probability of also getting ‘negative’ genes on both members of a chromosome pair is especially high. Inbred plants have little genetic diversity among each other. This allows for retaining genetics when in a controlled field environment like a seedstock increase isolation where all plants have identical, homozygous genes. Unfortunately, some of the affect is negative affect of some genes affecting plant size and perhaps some specific traits such as resistance to a pathogen. It is the corn breeders objective to match inbreds that will have dominant genes to reduce the negative affect when crossed with a specific inbred. After identifying the pair of inbreds that make desired hybrids with maximum expression of desired traits a commercial hybrid can be produced. Controlled pollination of one parent onto the other parent is essential for resulting seed to uniformly express the hybrid genetics. Few pollen from the female plant landing on its own ear silk, will result in seed with the female inbred and not that of the hybrid genetics. Such ‘self’ will be small, unproductive plants in the hybrid field. If female silks are pollinated by the wrong plants, such as from other hybrid fields, resulting seed will not be identical to the desired hybrid. Pollen produced by hybrid plants are not genetically identical to each other and therefore such contaminating pollen on a female plant in a hybrid seed production field results in seed with wide variation. Each outcross plant from this type of contamination will be different from the desired hybrid but also from each other, varying in plant height, ear placement and size and other noticed traits. Corn diversity benefits availability of traits and needs to be carefully controlled to maximize its advantages. Zea mays is a species that has sufficient morphological distance between the male and female flowers that most pollen does not fertilize its mother plant. This character is unique from most other crop plants in which the stamens and pistils are parts of the same flower. Although other species may have other strategies to reduce self pollination, like timing, the distance between the ear shoot and the tassel plus abundance of pollen that easily floats in air encourages pollination of other corn plants rather than self pollination.
This characteristic of corn promoted significant genetic diversity ultimately resulting in ease of adaptation to a wide range of environments as humans selected the genetics suited to their needs and environments. This is diversity to us as we selected genes for less starch in sweet corn, endosperm explosiveness in popcorn, soft starch in floury corn and hard endosperm of flint corn. Corn varieties are adapted to short seasons of temperate zones and to longer seasons near the equator. Even as pathogens evolve adaptation of some corn genetically controlled resistance, genetic variability within the species is eventually identified to produce the metabolic products needed to resist the pathogen. Open pollinated varieties maintain genetic heterozygosity as meiosis after fusion of male gamete with that of the egg cell, creates new mixes of the genes. Consequently, each seed in an open pollenated corn pollinated plant when at least one of the parents is heterozygous will have some genetic variation from adjacent seeds. Corn breeders have used this principle to get new, desired genetic characters. Genetic variability is advantageous when identifying useful characters, but not when attempting to get optimum grain production. The advantage of crossing two homozygous plants of two distinct genetic backgrounds is producing predictable and consistent hybrid plants. Controlled selfed pollination for several generation ultimately results in homozygosity, with essentially each seed having identical genetics. This selfing process also creates homozygosity of some genes that have negative affects on the plants, including characters affecting seed size and volume. Identifying homozygous inbreds that essentially overcome these negative genes of another hybrid parent is the plant breeders job for selecting potential high performing hybrids. Cross pollination characteristic of corn has allowed genetic variability and human effort to select and self pollinate to create inbreds and then crosses to make specific hybrids, allowing this species to be a major international crop. As many humans on earth face current interactions with a deadly virus, it is a reminder of significance of genetics and environmental interactions that affects our lives. Our genetic complexity affects each individual’s ability to defend against the Covid-19 virus but even that is qualified by the multiple individuals medical conditions because of past ‘environments’ and probably genetic changes in the relatively simple virus genes.
Human genetic diversity exists because of our evolution under multiple selection pressures of our past environments. All humans, despite evident, minor visible trait differences such as skin, hair and eye color, have other less evident genetic differences witnessed only by slight differences among an individual family. Corn has a genetic history somewhat similar to humans. Its ancestor was a related species and perhaps some crossing with another species but with human’s intervention it was moved to multiple environments and selection pressures allowing diversity of characters we could easily see. Yet, within each environment there was selection for other favored, less easily seen, characteristics. Just as with human the vast majority of genetics remain close to being identical to others of the species. Corn genetics that were best for desired traits in the season of an individual field in 2020 probably will not be the same as the best hybrid in another field nor even the best for the same farm in 2021. Small differences in 2021 environments will favor slightly different genetics affecting expression of traits not noted in 2020. Some traits observed in 2020 environments may not be expressed in all environments. We are dependent on maintaining genetic diversity within the corn species because of environmental variability. Humans also are dependent upon diversity not for those relatively simply inherited characters such as skin, hair and eye color but also for those less obvious genetics that our species successful adaptation to a forever changing environment. Success of our species, and of corn, is dependent upon genetic diversity. The biology of both species favors continual genetic interactions and diversity. 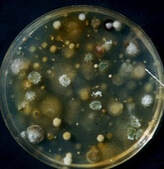 Those occupants of dying stalks, roots and ears will be there next spring as they continue to look for nutrition on organic material to digest. Living plants and animal tissue evolved with resistance to most of these organisms but we are all dependent on these organisms to release the components to be used again in the new generation.
Seeds are surrounded by many microorganisms mostly feeding on organic matter that is mostly dead. The photo, copied from our publication of Stalk Rot of Corn power point, shows the multiple colonies grown from a particle of soil in the spring of a corn field. These organisms are competing for the carbs locked up in the dead debris in the soil. Fusarium species are among these that can also penetrate some living corn roots. Fusarium is a fungus genus composed of many species. This group of fungi asexually produce spores called conidia that move in the wind. Most of these species also have a sexual stage in which the haploid hyphae fuse to form a diploid structure (ascus) in which the nuclei undergo meiosis, producing haploid ascospores, to produce more hyphae with new genetic combinations. The fact that the asexual and sexual stages are separate in time and, sometimes, location adds to the difficulty for specialists to associate the two. The ‘rules’ of naming these fungi declares that the sexual stage is primary. Consequently, on fungus commonly infecting corn seedlings, stalks and ears is known as Fusarium graminearum but also by its sexual stage name of Gibberella zeae, another common Fusarium infecting seedlings, stalks and ears is Fusarium verticillioides also known for the sexual stage name of Gibberella fujikuroi. Adding to the naming confusion, the previous name for F. verticillioides was Fusarium moniliforme. The names are confusing, but biology of these fungi is not much better. They tend to mostly be weak pathogens. They can enter weakened root and mesocotyl tissue of the germinating seed. Host resistance seems to limit the fungus ability to destroy much tissue, but it appears to simply live in the corn tissue without causing visible damage. It is not unusual to find at least a few seed germinating on a paper towel that look normal but have some hyphae of Fusarium verticillioides growth. This species frequently can be isolated from corn leaf tissue with no apparent damage. Likewise, Fusarium species are nearly always isolated from stalks of corn plants. Often, if no symptom associated with another pathogen, such as the black streaks in outer rind caused by the anthracnose fungus (Colletotrichum graminicola), we tend to call it Fusarium stalk rot. The death of the stalk mostly was a biological problem of the corn plant and fungi like Fusarium was there, among others, to assist in the digestion of dead stalk tissue. Fusarium is the name of genus of a fungus group of species that are common saprophytes and plant pathogens. Ability to kill living cells varies among Fusarium species but certainly they can feed on dead cell tissue. Among the many fungal species feeding on dead corn stalks almost always a Fusarium species will be present. If no diagnostic indicators of Gibberella, anthracnose or Diplodia are present, it becomes easy to call the disease Fusarium stalk rot.
Fusarium species that are often found in dead and live corn plant stalks initially grow between cells, producing enzymes to digest the walls of the pith cells and fibers. These walls are composed of complex cellulose compounds combined with proteins and basic carbohydrate molecules, formed by biological activity within the living cells and secreted through the cellular membranes. These compounds protect the cellular contents by structure and some active resistance products as well. Cell wall components include cellulose, arabinoxylans, hydroxycinnamates, pectins, glycoproteins and lignins. Fungi initially grow between the cells, secreting enzymes to digest the cell wall structures. Meanwhile living corn cells produce compounds such as Jasmonic acid, abscisic acid and salicylic acid to limit the growth of the Fusarium species. This battle begins with the first incursion of the fungus, perhaps with injury to the seedlings or perhaps with corn borer. Artificially infecting the stalk with any of the fungi associated with corn stalk rot shortly after pollination usually shows infection limited to area near to point of inoculation. As the tissue matures the fungi seem to more rapidly spread, digesting more cell walls. Breaking down the complex compounds allows the fungus to utilize more simple carbohydrates for their own metabolism. Digestion of the cell wall components weakens the strength of the stalk beyond the sudden decrease of strength that occurred with the plant wilted after root rot. The wilting caused the pith cells to withdraw from the rind, effectively changing the structure from a rod to a tube. This alone is believed to reduce strength by one-third. Corn hybrids that tend to construct more and stronger cells in the rind can have less lodging but the physical change from a rod to tube is difficult to match by cell wall strength alone. Maintaining sturdy stalks until harvest is a balance among producing sufficient carbohydrates for filling all kernels, avoiding wilt by delaying death of root cells and maintaining stalk cells alive to fend off potential pathogens such as Fusarium |
About Corn JournalThe purpose of this blog is to share perspectives of the biology of corn, its seed and diseases in a mix of technical and not so technical terms with all who are interested in this major crop. With more technical references to any of the topics easily available on the web with a search of key words, the blog will rarely cite references but will attempt to be accurate. Comments are welcome but will be screened before publishing. Comments and questions directed to the author by emails are encouraged.
Archives
December 2021
Categories
|
 RSS Feed
RSS Feed